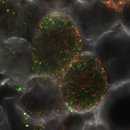
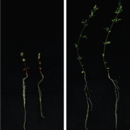
Symbiotic and pathogenic bacteria use common strategies to colonise plants, such as biofilm formation, quorum-sensing, hormone-balance manipulation and protein secretion, among others. However, in the majority of the cases pathogenic bacteria remain on the plant surface or in the apoplast and are not able to invade cells. Bacteria of the order rhizobiales are among the few that are able to colonise the interior of plant cells. This bacterial order is diverse and includes plant symbiotic bacteria (Mesorhizobium, Bradyrhizobium), plant pathogens (Agrobacterium, Candidatus Liberibacter) and animal pathogens (Brucella, Bartonella).
more